안녕하세요?
지난번 포스팅에서 TCR이 다양한 유전적인 이유에 대해서 포스팅할 차례라고 언급을 하고 나서 끝을 냈었는데, 이번 포스팅에서는 먼저 T 세포 수용체(TCR)의 유전자 재배열에 대해서 포스팅을 하고나서, NK수용체에 대한 내용을 추가적으로 다루고자 합니다. 먼저 간단하게 αβ TCR에 대해서 다루겠습니다.


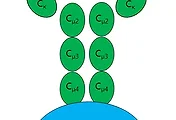
먼저 위 그림에 묘사가 된 것처럼 TCR을 구성하는 α사슬은 14번 염색체에 유전자 정보가 위치하고, β사슬을 구성하는 유전자는 7번 염색체에 위치를 하고 있습니다. 어디서 한번은 본적이 있다는 생각이 든다면 맞는 것이, 항체의 V(D)J recombination과 같이 흉선에서 T세포가 분화되어 나올 때 이러한 유전자의 재조합이 일어나고, 이어서 TCR의 발현까지 발현까지 영향을 주게 됩니다.
링크 : 항체가 다양한 이유-유전자상에서 일어나는 이야기 part2
위 링크에서 다룬적이 있었는 RAG복합체와 DNA변형 효소가 여기서도 동일하게 작용한다고 면역학 책에서 언급되어 있습니다. B세포와 마찬가지로 T세포 역시 V(D)J recombination이라는 동일한 과정을 이용해서 항체가 다양한 것처럼, T세포 표면 수용체(TCR)도 다양하게 만들 수 있는 것입니다.

위 그림은 β사슬의 유전자 재조합 과정에서 '기하학적 재조합(geometric recombination)' 이라는 것이 일어난다는 것을 보여주고 있습니다. 일반적으로 Dβ1이 사용되었다면, 한 세트(?)인 Jβ1시리즈가 올 것 같지만, 다음 세트(?)에 있는 Jβ2.2가 오는 식으로 생각지도 못하게 다양한 조합을 만들어 낼 수 있는 것입니다. 하지만 이것이 끝이 아닙니다.

예전에 항체의 '유전적 다양성'에 대해서 포스팅을 할 때 언급이 되었는 '이음부 다양성(junctional diversity)'가 여기서도 적응이 됩니다. 위 그림에서 묘사가 된 것처럼 V,D,J가 '가변 경계 재조합(variable boundary recombination)'으로 인해서 여러가지 '접합부 서열(junctional sequence)'가 나오게 됩니다. 즉, 접합부에도 이런 변화가 생겨서 다양한 형태의 TCR이 나오게 됩니다.

여기서 일전에 설명을 드린 γδ TCR에 대해서 궁금하실 건데요, 위 그림에서 δ사슬이 염색체 14번에서 어떻게 위치하고 있는지를 나타낸 그림입니다. 사람의 δ사슬은 14번 염색체의 Vα와 Jα 사이에 위치하고 있으며, 유전자가 재조합시에 필연적으로 δ사슬 좌위의 불활성화를 일으킨다고 합니다. 즉, 대다수의 경우에는 α사슬이 발현될 경우 δ사슬의 유전자는 자동으로 떨어져 나간다는 의미입니다.

반대로 7번 염색체에 γ사슬의 유전자가 위치해 있기는 하지만 δ사슬과는 다르게 β사슬의 유전자 좌위와는 아무런 관계가 없이 위치해 있다는 것이 문제입니다. 면역학 책에 서는 아직 γδ TCR에 대해서 그다지 규명된 것은 많지 않으나, '상피조직'에서는 우세한 T세포 집단이 될 수 있다고 합니다.
이제 NK세포(자연 살해 세포)의 수용체라고 해야 할까요? 이 NK세포에 관련이 되어 있는 일에 대해서 깊이 들어가 봐야 겠습니다. 예전 포스팅에서는 언급하기를, NK cell이 이미 '감염된 세포'를 자살시킨다고 하였습니다.
링크 : 선천적 or 내재적 면역 이야기 final-선천적 면역의 마지막 이야기
이 NK세포가 건강한 신체 조직을 공격해서 죽이면 안되기에, NK세포 활성 수용체 외에도 자실들의 기능을 억제하는 수용체 역시 가지고 있다고 합니다. 이 수용체들은 '내장된(hard-wired)' 수용체라고 해서, 다양성을 위해서 VDJ 재조합에 들어가지는 않는다고 합니다. 그래서 면역학 책에서는 '선천적 or 내재적' 면역 반응에서 언급된 적이 있는 '패턴인식 수용체'와 유사하다고 언급이 되어 있습니다.

위 그림은 정상적으로 자기 MHC class I를 발현하는 인체의 체세포와 만났을 때, NK 세포가 '무반응'하는 것을 묘사한 그림입니다. 그럼 이 MHC class I가 무엇이냐는 의문이 드실 것인데, 먼저 아래의 링크를 타고 내용을 읽어 주시기 바랍니다.
링크 : 적응면역 이야기 part3-T cell 이야기 part1
위 링크에 나온 내용에 의하면, 병원체에 '감염'되어 숙주가 된 인체의 세포는 MHC class I를 통해서 '세포독성 T 세포'를 끌여들이게 됩니다. 따라서 미생물들은 이 MHC class I을 아예 발현되지 못하게 만드는 전략을 취하기도 하는데, 이럴 경우 아래와 같은 그림의 상황이 벌어지게 됩니다.

위 그림에서 체세포는 바이러스와 같은 '항원'에 감염이 되어서 MHC class I이 전혀 발현이 되지 않은 상황입니다. 이 상황에서 '감염된' 체세포에서 '활성 리간드'까지 발현이 되면, NK세포가 망설일 것도 더 없이 표적이 된 세포를 공격해서 세포 자살이라는 apoptosis를 일으키게 되는 것입니다.
그런데 위 그림들에서 묘사한 NK세포들의 작용에 대한 것도 '가설'이라고 해서, MHC class I가 존재하면서도, NK 세포를 활성화 시키는 리간드가 존재하는 '모순'적인 상황이 발생하게 됩니다.

위 그림처럼 MHC class I와 활성 리간드가 동시에 존재할 경우에는 '억제'하는 신호의 강도와 '활성화'하는 신호의 강도 사이에서 어느쪽 신호가 더 강하느냐에 따라서, NK세포가 활성화 되어서 체세포를 '죽게'만들거나, 아니면 억제가 되어서 아무런 작용도 하지 않을 것입니다.

마지막으로 위 그림처럼 MHC class I와 활성 리간드 2개 모두 다 동시에 없는 경우 입니다. 이런 경우라면 NK세포가 반응을 아예 할 수 없어서, 면역학 책에서는 거의 '인식할 수 없는 (ignore)'라고 언급하고 있습니다. 이것으로 NK세포 매개 살해와 'missing-self'가설에 대해서 대략적으로 설명하였습니다.
원래라면 여기서부터 NK세포의 활성화 및 억제 수용체에 대해서 더 나아가야 하지만, 예상과는 달리 NK수용체에 대한 내용도 많아서 포스팅을 더 이어나가면 너무 길어지는 문제점이 있어 보입니다. 그래서 이번 포스팅은 이쯤에서 마치도록 하고, 다음 <면역학 이야기> 포스팅에서는 나머지 부분을 다루도록 하겠습니다. 긴글 읽어 주셔서 감사합니다.
'생명과학 이야기 > 면역학 이야기' 카테고리의 다른 글
| T세포 표면 수용체(TCR)에 대한 상세한 이야기 part1 (33) | 2017.09.17 |
|---|---|
| B세포 표면 수용체(BCR)에 대한 세부적인 이야기 part2 (0) | 2017.09.11 |
| B세포 표면 수용체(BCR)에 대한 세부적인 이야기 part1 (4) | 2017.09.08 |
| 항체가 다양한 이유-유전자 상에서 일어나는 이야기 part3 (0) | 2017.09.04 |
| 항체가 다양한 이유-유전자상에서 일어나는 이야기 part2 (0) | 2017.09.01 |