안녕하세요?
지난번 포스팅에서 약속드린 대로, 이번 포스팅에서는 면역 글로불린이라고 불리는 항체의 5가지 모양에 대해서 대략적으로-면역학책에서 이 이상은 들어가지 않은 부분은 저로서도 방법이 없습니다- 들어가 보고, 어떻게 항체의 다양성이 나오는 지에 대해서 V(D)J 재조합에 대해서 포스팅을 하고자 합니다. 먼저 항체의 5가지 변형이라고 해야 할까요? 5가지 종류 중에 하나인 IgM에 대해서 알아보겠습니다.

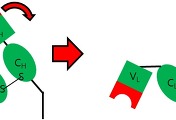
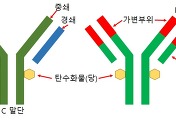
위 그림은 IgM의 단량체 단위로, 실제로는 이렇게 존재하지를 않고, 5량체로 합체한 상태로 혈청 내에서 존재하고 있다고 합니다. 위 그림에서는 지난번 포스팅에서 다룬 '경첩'을 지닌 것처럼 묘사가 되었지만, 경첩부위에 하나의 '일정부위'를 더 보유하는 것으로 IgG와는 구별이 되는 차이점이라고 합니다. 그래서 간략하게 다시 나타내면 아래의 그림과 같이 된다고 합니다.

위 그림에서 IgM은 4개의 CH도메인으로 이루어져 있는 형태로 위 그림처럼 간략하게 나타낼 수 있는 것입니다. 그럼 아까전에도 설명을 했다시피 '오량체'라고 해서 5개의 조각이 합쳐져서 존재를 하는데, 연결 사슬이라는 의미에서 'J사슬'이라고 불리우는 '폴리펩티드'에 의해서 연결되어 있다고 합니다. 아래의 그림은 간략하게 IgM의 오량체 구조를 묘사한 그림입니다.

이렇듯 IgM은 5량체로 혈액속에 존재하고 있으며, 면역학책에 의하면 10개의 항원결합부위를 가지고 있기 때문에 병원체의 표면에 강력하게 결합을 한다고 합니다. 이와는 비슷하지만 다른 것이 IgA로, IgA는 J사슬의 도움을 받아서 이량체로 존재를 할 수도 있지만, 림프절, 비장, 골수등의 형질세포에 의해서 '단량체'로 만들어져 분비됩니다.

위 그림은 단량체로 분비가 된 IgA의 구조를 나타낸 그림입니다. 일단 IgG1과 면역학 책에서 언급하기를 닮았다고 하기는 하지만, O-glycosylation이라고 해서 N-glycosylation과는 다른 형태의 당을 가지면서 상대적으로 긴 형태의 경첩을 가지고 있다고 합니다. 혹시 이 O-glycosylation이니, N-glycosylation이니 하는 단어가 생소하신 분들은 아래의 링크를 읽어 주시기 바랍니다.
링크 : 단백질의 당화(Protein Glycosylation)
그럼 반대로 이합체가 된 IgA는 주로 점막 상피에 있는 면역 조직에서 만들어지며, 주로 창자내로 분비가 된다고 합니다. 이외에도 젖, 침, 땀, 눈물 등의 분비물에 주로 존재하는 항체라고 합니다. 아래의 그림은 간단하게 이량체인 IgA를 묘사한 그림이 되겠습니다.

IgM의 경우처럼, J사슬이라는 것이 있어서 2개의 단일 IgA가 연결이 된 형태로 만들어 줍니다. 그리고 나서 끝인가 하면 그것도 아닌것이, 표피세포로 부터 분비되기 위해서는 '분비성 구성요소'라고 하는 일종의 펩타이드 조각이 붙게 됩니다. 왜 이런 구성 요소가 붙는가 하면, 아래의 그림과 같은 작용이 일어나기 때문이라고 합니다.

위 그림에서 대해서 간략하게 설명을 하자면, 우선 표피세포는 기저막(basal membrane)이라 해서, 인체의 내부쪽으로 면역 글로불린(Immunoglobulin, IG)를 잡을 수 있는-정확히는 Poly Ig를 잡을 수 있는 수용체(receptor)를 만듧니다. 이게 바로 위 그림에서 표시가 된 Poly Ig receptor라는 것이며, 여기다가 이량체 IgA가 결합하면 표시세포 내에 있는 endocytic vacuule을 통해서 점막 표면(apical surface)로 수송이 된다고 합니다.
마자막 단계에 이르러서는 Poly Ig receptor의 일부를 잘라서 분비를 하게 되는데, 이와 같은 과정을 위 그림에서 가위와 절취선으로 묘사를 하였습니다. 일단 수용체의 일부를 끊으면, IgA와 수용체가 붙은 채로 점막 표면으로 분비가 되게 되는데, 이때 이량체인 IgA에 붙어 있는 수용체의 단편을 '분비성 구성요소(secretory component)'라고 하며, 전체 분자를 가르켜서 '분비성 IgA(secretory IgA)'라고 한다고 합니다.

마지막으로 다루게 될 것이 IgE와 IgD입니다. 우선 IgE는 특별한 것은 없고, IgM의 단량체 단위와 유사하다고 면역학 책에서 언급이 되어 있습니다. IgD의 경우에는 경첩이 상당히 길어서, O-glycosylation이 풍부한 부위와 전하가 풍부하며 나선형으로 아미노산이 꼬여진 구조를 형성하는 부위로 나누어 질 수 있습니다. 다만 단백질 분해 공격에 IgD는 취약하다는 단점이 있습니다.
예전에 항체가 다양한 이유로 V(D)J 재조합에 대해서 언급을 하였습니다만, 그 때는 자세한 설명 없이 넘어 갔었습니다. 그러나 이번 포스팅에서는 어떻게 항체가 유전적으로 다양성을 확보할 수 있는지에 대해서 들어가 볼 시간이 되었습니다. 조금은 어려울 수가 있지만, 그대로 이해하기 쉽도록 풀어서 서술을 하도록 하겠습니다.
먼저 항체(antibody)의 유전정보는 1개의 염색체 안에 완전히 존재하는 것이 아니라, 여러개의 염색체에 나누어져 있다는 사실을 언급해야 겠습니다. 이걸 무엇에 비유해야 할까요? 전투기를 만들어야 하는데, 설계도인 청사진이 온전히 공장 한군데게 보관되어 있지 않고, 직소퍼즐마냥 흩어져서 여러 공장에 나누어져 보관되어 있다고 해야 할듯 합니다.

위 그림에서 표시가 된 염색체인 2번, 14번 22번에 면역 글로불린을 형성하는 유전자가 각각 위치하고 있다고 합니다. 그리고 더 복잡하게도 염색체 하나에 1개의 유전자로 또 완전하게 있지를 않고, 잘게 나누어져 있다고 합니다. 무슨 말인지 당장은 이해하기 어려우신 분들을 위해서 간략하게나마 그림으로 묘사를 할까 합니다. 먼저 아래의 그림을 봐 주시기 바랍니다.

위 그림에서 묘사가 된 것처럼, 우리가 흔히 아는 IgG의 중쇄와 경쇄는 1개의 염색체가 아니라, 중쇄는 14번 염색체에 정보가 있고, 경쇄의 λ(람다)와 Κ(카파)부위는 각각 22번과 2번 염색체에 있습니다. 그리고 이 염색체들 안에도 하나의 온전한 DNA서열로 존재하는 것이 아니라 마치 레고블럭처럼 여러개로 나누어진 형태로 존재를 하고 있다고 합니다.
원래라면 여기서 본격적으로 이 레고 블록에 대한 설명에 들어가야만 합니다만, 그러기에는 포스팅이 너무 길어지는 감이 있기에 여기서 마치도록 하겠습니다. 면역학이 어렵기는 하지만, 관심을 가지고 여기까지 들어주신 분들에게 감사를 표하면서, 혹시나 잘못된 부분에 대해서는 댓글을 달아 주시면 감사하겠습니다.
'생명과학 이야기 > 면역학 이야기' 카테고리의 다른 글
| 항체가 다양한 이유-유전자상에서 일어나는 이야기 part2 (0) | 2017.09.01 |
|---|---|
| 항체가 다양한 이유-유전자상에서 일어나는 이야기 part1 (4) | 2017.08.29 |
| 항체에 대한 세부적인 이야기 part2 (0) | 2017.08.23 |
| 항체에 대한 세부적인 이야기 part1 (8) | 2017.08.20 |
| 적응면역 이야기part4-T cell이야기 part2 (0) | 2017.08.15 |